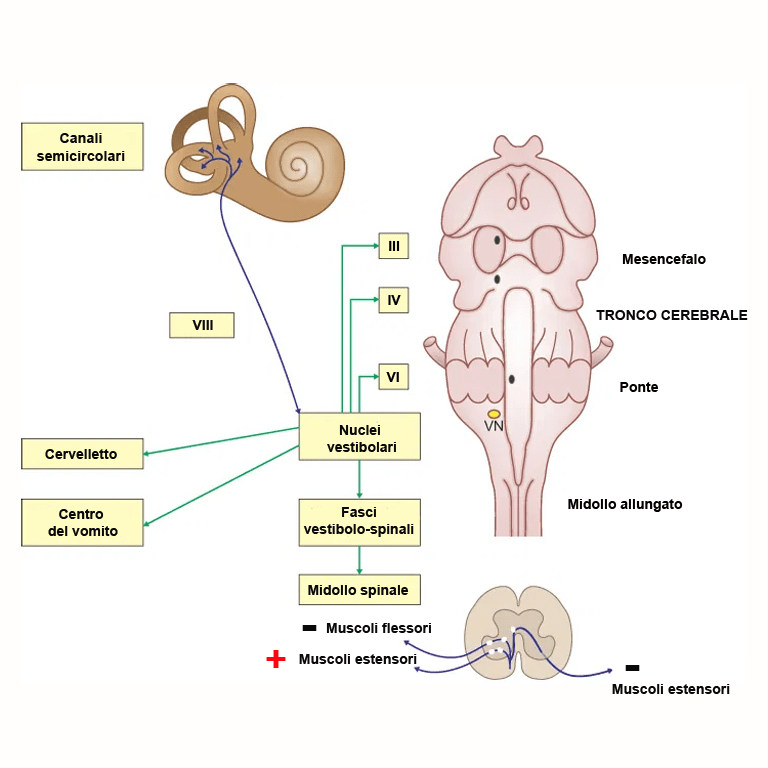
Nervi cranici che controllano i movimenti oculari e riflessi vestibolari
Il III (n. oculomotore), IV (n. trocleare) e il VI paio (n. oculomotore) di nervi cranici controllano i movimenti oculari. Tutti questi nervi sono formati da fibre motrici somatiche che si distribuiscono ai mm. estrinseci dell’occhio. Il n. oculomotore è anche costituito da fibre motrici viscerali parasimpatiche. Le fibre motrici somatiche dei tre nervi originano da neuroni situati nel tegmento del tronco cerebrale e raggruppati in tre diversi nuclei motori somatici I nuclei motori somatici del III e del IV si trovano entrambi nel mesencefalo. Naturalmente (dal momento che i nervi cranici sono numerati progressivamente in senso rostro-caudale) quello del III è il più rostrale. Il nucleo motore somatico del VI si trova, invece, nel ponte. Il n. trocleare innerva esclusivamente il m. obliquo superiore dell’occhio, il n. abducente innerva il m. retto mediale dell’occhio e il m. retrattore dell’occhio. I rimanenti muscoli estrinseci dell’occhio (m. retto dorsale, ventrale e laterale e il m. obliquo inferiore) sono innervati dal n. oculomotore.
Attraverso le fibre cortico-nucleari che raggiungono i nuclei motori di questi nervi il fascio cortico-spinale controlla i movimenti volontari degli occhi quando l’animale fissa lo sguardo in un’area specifica del proprio campo visivo. Gli occhi però si muovono anche con movimenti riflessi che sono attivati da fibre dei nuclei vestibolari situati nel tegmento del ponte.
Le proiezioni dai nuclei vestibolari si estendono a diversi nuuclei di neuroni del tronco cerebrale, al midollo spinale e al cervelletto. I movimenti oculari riflessi (riflessi vestibolo-oculari) dipendono dalle fibre che terminano nei nuclei motori dei nervi oculomotore, trocleare e abducente e intervengono nel determinare i movimenti coniugati degli occhi per fissare lo sguardo su un determinato bersaglio. Questi circuiti fanno capo al labirinto dinamico dell’orecchio interno. Alcuni degli assoni che originano dal nucleo vestibolare laterale (il più sviluppato) e dai nuclei vestibolare mediale e caudale ricevono afferenze dalle macule (labirinto statico) vanno a costituire i fasci vestibolo-spinale laterale e vestibolo-spinale mediale che terminano facendo sinapsi sui motoneuroni spinali che innervano i muscoli del collo, del tronco e degli arti. In particolare, la stimolazione dei nuclei vestibolari aumenta il tono dei muscoli estensori (antigravitari) e dei muscoli flessori (gravitari) omolateralmente e inibisce il tono muscoli estensori controlaterali. Questi circuiti riflessi (riflessi vestibolo-spinali) sono fondamentali nel mantenimento dell’equilibrio sia in stazione che durante il movimento. Le proiezioni al cervelletto consentono la coordinazione della posizione degli occhi, del collo, del tronco e degli arti con la posizione della testa (propriocezione inconscia). Inoltre, le proiezioni alla corteccia cerebrale consentono la percezione cosciente del movimento e della gravità (propriocezione conscia) necessaria a una corretta esecuzione dei movimenti volontari.
Altri assoni provenienti dai nuclei vestibolari raggiungono il centro emetico della formazione reticolare. Questo spiega perché in alcune patologie che colpiscono il sistema vestibolare può manifestarsi vomito.

Se vuoi saperne di più e capire meglio scorri la pagina verso il basso
Il sistema vestibolare
Il sistema vestibolare è quell’insieme di strutture che, a partire da recettori localizzati nel vestibolo dell’orecchio interno, raccoglie e elabora informazioni sull’equilibrio del corpo nello spazio. Le componenti anatomiche del sistema vestibolare sono suddivise in periferiche e centrali.
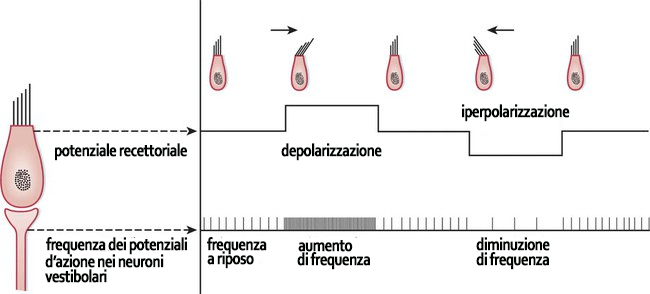
Le componenti periferiche sono contenute nella porzione petrosa dell’osso temporale (rocca petrosa o osso petroso) in cui è contenuto l’orecchio interno. Quest’ultimo è formato da un complicato sistema di canali e spazi ossei scavarti all’interno dell’osso petroso, il labirinto osseo, entro cui è contenuto un altrettanto complicato insieme di formazioni a parete sottile, membranosa, il labirinto membranoso, che ne ricalca la forma. Entrambi i labirinti sono costituiti da un vestibolo (osseo e membranoso) e da una coclea (chiocciola – ossea e membranosa) e sono separati fra loro da uno spazio ripieno di perilinfa, un liquido con composizione simile a quella dei liquidi extracellulari. All’interno del labirinto membranoso è, invece, contenuta l’endolinfa, un liquido con caratteristiche simili al citosol. Mentre nel vestibolo membranoso sono localizzati i recettori dell’equilibrio, nella coclea membranosa sono localizzati i recettori acustici. I recettori dell’equilibrio, che fanno parte della componente periferica del sistema vestibolare, si trovano nelle macule, a livello del sacculo e dell’utricolo, e nelle creste ampollari, a livello dei canali semicircolari. La componente periferica del sistema vestibolare è completata dai neuroni del ganglio vestibolare che, con i loro dendriti, formano giunzioni cito-neurali con le cellule capellute, i recettori statocinetici vestibolari, e, con i loro assoni, costituiscono il n. vestibolare che termina nel ponte. I recettori statocinetici delle macule e delle creste ampollari sono cellule sensitive secondarie. Si tratta di cellule epiteliali modificate che si sono specializzate nella trasduzione di segnali meccanici (movimenti dell’endolinfa) in segnali elettrici. Il loro polo apicale è modificato in maniera caratteristica: presenta lunghe stereociglia e un chinociglio molto sviluppato, motivi per cui queste cellule sono chiamate cellule capellute. A riposo, quando le stereociglia non sono deviate, i neuroni sensitivi primari del ganglio vestibolare, che fanno sinapsi al polo basale delle cellule capellute trasmettono spontaneamente circa 100 potenziali d’azione al secondo. Quando le stereociglia si piegano in direzione del chinociglio, le cellule si depolarizzano e rilasciando il loro neurotrasmettitore (glutamato o aspartato) alle giunzioni cito-neurali con i neuroni vestibolari, che, di conseguenza, aumentano la frequenza dei potenziali d’azione. Quando le stereociglia sono piegate in direzione opposta, le cellule capellute si iperpolarizzano, il rilascio del trasmettitore diminuisce e la frequenza del potenziale d’azione dei neuroni sensoriali diminuisce. Pertanto, lo spostamento delle stereociglia delle cellule capellute in una di queste direzioni può essere rilevato dal cervello come un aumento o una diminuzione della frequenza del potenziale d’azione a riposo. Le deviazioni in altre direzioni sono molto meno efficaci.
Le cellule capellute delle macule registrano i movimenti di accelerazione lineare della testa lungo il piano orizzontale (nel quale è orientata la macula dell’utricolo) o verticale (nel quale è orientata la macula del sacculo). Nell’insieme costituiscono quello che funzionalmente si definisce labirinto statico dell’orecchio interno. Gli statocettori delle macule sono ricoperte da una matrice gelatinosa in cui sono incorporati piccoli corpi cristallini (otoliti). Gli otoliti esercitano una trazione sulle stereociglia delle cellule capellute in risposta alla gravità. Di conseguenza, le stereociglia si piegano e questo stimola il nervo vestibolare, come descritto sopra. Così le macule rispondono all’accelerazione lineare della testa, sul piano orizzontale (mentre l’animale cammina o corre, per esempio) o sul piano verticale (con l’accelerazione di gravità). La funzione principale del labirinto statico è quella di indicare la posizione della testa rispetto al tronco. In risposta a questo segnale, i nuclei vestibolare laterale e mediale attivano movimenti compensatori per l’equilibrio (riflessi vestibolo-spinali). Questi movimenti hanno l’effetto di mantenere il centro di gravità entro la base d’appoggio quando l’animale è in stazione (equilibrio statico) o immediatamente davanti alla base di appoggio durante la locomozione (equilibrio dinamico). Inoltre, consentono alla testa di rimanere verticale. Il labirinto statico controlla l’equilibrio tramite i fasci vestibolo-spinali laterale e mediale. Esso funziona in associazione con i propriocettori della muscolatura degli arti e i fotorecettori della retina, per il mantenimento della postura eretta. Il labirinto statico contribuisce anche alla consapevolezza segmentale: cioè, il senso di conoscere la posizione del corpo nello spazio. Questo è un vero e proprio sesto senso, normalmente fornito da tre sistemi: il sistema visivo, il sistema propriocettivo e il sistema vestibolare. Se privato di uno di questi sistemi, l’animale è comunque in grado di rimanere in stazione e muoversi utilizzando la consapevolezza fornita dagli altri due sistemi. Ad esempio, dopo la perdita della vista, un animale può ancora camminare e rimanere in posizione verticale, nonostante i vincoli della cecità. Dopo la perdita della propriocezione cosciente, gli animali utilizzano la vista come sostituto. In queste circostanze, privare un animale degli stimoli visivi (coprendogli gli occhio rende atassico (atassia sensoriale). Allo stesso modo, se i labirinti statici sono inattivi a seguito di un qualsiasi tipo di lesione, la semplice copertura degli occhi potrebbe causare una caduta (atassia vestibolare).

Rifletti sul fatto che il neurologo può effettuare test clinici che sfruttano queste nozioni per diagnosticare correttamente i diversi tipi di atassia.
I tre canali semicircolari membranosi si trovano all’interno dei canali semicircolari corrispondenti di ciascun labirinto osseo. I recettori in essi contenuti formano il labirinto dinamico o cinematico. I canali semicircolari sono orientati approssimativamente ad angolo retto tra loro, lungo le tre direzioni dello spazio, ed entrambe le estremità di ciascun condotto, riempito di endolinfa, terminano nell’utricolo, in prossimità del quale si osserva una dilatazione chiamata ampolla. Ciascun’ampolla contiene una cresta ampollare dove si localizzano le cellule capellute le cui stereociglia sono incorporate in una massa gelatinosa (cupola) che si fissa al tetto dell’ampolla. Tutte le cellule capellute di una data cresta ampollare sono orientate nella stessa direzione rispetto alle loro stereociglia. Insieme, la cresta delle cellule capellute e la cupola sovrastante occupano l’intero diametro dell’ampolla. Il movimento dell’endolinfa nei canali semicircolari provoca la deflessione della cupola, che è orientata trasversalmente alla direzione di flusso. Questa deflessione piega la stereociglia dando l’avvio alla trasduzione del movimento in segnale elettrico. Poiché i tre canali semicircolari sono perpendicolari tra loro, il movimento della testa in qualsiasi piano o una sua rotazione angolare agisce su una cresta ampollare e stimola i neuroni vestibolari. Le creste funzionano in equilibrio dinamico fra loro e sono coinvolti nella trasduzione dell’accelerazione e della decelerazione angolare della testa. Quando la testa inizia ad accelerare con un moto rotatorio, il canale semicircolare e la cresta ruotano con la testa, ma l’endolinfa contenuta all’interno si muove più lentamente per inerzia. Questa differenza relativa nella velocità di accelerazione del canale semicircolare e dell’endolinfa in esso contenuta fa sì che la cresta collida con l’endolinfa che si muove più lentamente. Ciò determina uno spostamento della cupola in direzione opposta a quella di rotazione della testa con una corrispondente flessione delle stereociglia delle cellule capellute. L’opposto accade con la decelerazione perché il canale semicircolare e la cresta rallentano immediatamente insieme alla testa mentre l’inerzia continua a spingere in avanti l’endolinfa. La stimolazione della cresta avviene soltanto l’accelerazione o la decelerazione rotatoria della testa , non durante la velocità di rotazione costante. Quando la velocità di rotazione è costante, il movimento dell’endolinfa alla fine eguaglierà il movimento dei dotti semicircolari e le stereociglia non saranno più piegate. I canali semicircolari situati sui lati opposti della testa, ma approssimativamente sullo stesso piano (complanari), lavorano in coppia per fornire al cervello informazioni sulla direzione e la natura del movimento della testa. Ad esempio, un’accelerazione rotatoria in senso orario della testa provocherebbe la flessione delle ciglia delle cellule capellute sensibili alla direzione in ciascun membro di una coppia complanare di condotti semicircolari sui lati opposti della testa. Tuttavia, le cellule capellute della cresta sul lato destro della testa (corrispondente alla direzione del movimento) si depolarizzano con aumento della frequenza dei potenziali d’azione nel n. vestibolare omolaterale, mentre quelle del condotto del lato sinistro si iperpolarizzano con riduzione della frequenza dei potenziali d’azione nel n. vestibolare controlaterale alla direzione del movimento. L’encefalo interpreta i mutamenti reciproci nella frequenza del potenziale d’azione sensoriale come risultanti dall’accelerazione o decelerazione in senso orario o antiorario in un dato piano di movimento. In realtà, l’accelerazione/decelerazione rotativa in un dato piano di solito colpisce tutti e tre i set di canali semicircolari accoppiati, ma ogni coppia in gradi diversi. In questo modo, il sistema bilaterale di sei condotti semicircolari rileva la direzione sia dell’accelerazione che della decelerazione angolare della testa e attiva o inibisce determinate strutture del SNC per produrre la risposta riflessa appropriata, stabilizzando gli occhi (riflessi vestibolo-oculari) quando l’animale muove il capo.
I componenti centrali del sistema vestibolare sono costituiti dai nuclei vestibolari del ponte i cui neuroni inviano assoni al cervelletto, al midollo spinale e al tronco encefalico rostrale. Attraverso i fasci vestibolo-spinali, le proiezioni al midollo spinale hanno un effetto facilitante sui muscoli estensori (antigravitari) ipsilaterali dell’arto consentendo all’animale di mantenere l’equilibrio durante la stazione e la locomozione (riflessi vestibolo-spinali). Tramite il fascicolo longitudinale mediale, le proiezioni ai nuclei motori somatici dei nervi cranici che controllano i mm. oculari estrinseci (III, IV e VI) controllano i movimenti riflessi degli occhi (riflessi vestibolo-oculari). Questi ultimi sono fondamentali per mantenere le immagini immobili sulla retina. Le proiezioni verso il talamo e, di qui, alla corteccia cerebrale consentono la percezione cosciente della posizione, mentre quelle al cervelletto sono responsabili del mantenimento della coordinazione di occhi, collo, tronco e arti in relazione ai movimenti della testa. Infine, quelle alla formazione reticolare del tronco cerebrale raggiungono il centro responsabile del vomito e della cinetosi.
Prova a rispondere a queste domande:
- Cosa si intende per cellula sensitiva secondaria?
- Quali sono i neuroni di primo e di secondo ordine nel sistema vestibolare?
- I neuroni del nucleo vestibolare laterale sono di grosse dimensioni. Perchè questa caratteristica è importante dal punto di vista funzionale?
- Cosa si intende per atassia?
- Con quale parte del cervelletto sono colllegati i nuclei vestibolari?
(le risposte sono al fondo della pagina)

Ricordati di consultare un libro o cercare informazioni in rete se non conosci il significato di alcuni termini!
Risposte
1. Una cellula sensitiva secondaria è un recettore costituito da una cellula epiteliale modificata. Sono cellule sensitive secondarie le cellule capellute, le cellule acustiche e le cellule gustative. L’aggettivo “secondaria” è utilizzato per distinguerle dalle cellule sensitive primarie che sono, invece, neuroni come i neuroni sensitivi primari dei gangli spinali o dei gangli sensitivi associati ai nervi cranici, i fotorecettori della retina o i neuroni olfattivi.
2. I neuroni di primo ordine del sistema vestibolare sono i neuroni sensitivi primari del ganglio vestibolare, quelli di secondo ordine sono i neuroni dei diversi nuclei vestibolari. I termini si riferiscono alla sequenza con la quale gli stimoli sensitivi agiscono su questi neuroni lungo le diverse vie afferenti. Fai attenzione che i termini neuroni di primo ordine e neuroni sensitivi primari non sono sinonimi fra loro.
3. I neuroni del nucleo vestibolare laterale sono fra i più grossi neuroni del SNC. Essi danno origine al fascio vestibolo-spinale laterale costituito da assoni mielinici anch’essi di grosso diametro. Esiste, infatti, una relazione diretta fra le dimensioni del corpo cellulare di un neurone e quelle dell’assone che ne deriva. Queste caratteristiche sono importanti perché la velocità di conduzione dello stimolo nervoso è direttamente proporzionale al diametro della fibra nervosa lungo cui lo stimolo si muove. Per poter efficacemente innescare i riflessi vestibolo-spinali che consentono di evitare una caduta quando perde l’equilibrio occorre che la risposta sia estremamente veloce per far si che il baricentro non si sposti al di fuori della base d’appoggio, con compromissione dell’equilibrio dell’animale .
3. L’atassia (dal greco ataxiā, disordine) è un disturbo della coordinazione muscolare che rende difficoltosa e imprecisa l’esecuzione di alcuni movimenti volontari. Secondo le strutture responsabili, si distinguono diversi tipi di atassia: l’atassia motoria deriva da lesioni delle vie motrici somatiche, l’atassia sensoriale (o sensitiva) deriva da lesioni delle vie propriocettive, l’atassia vestibolare da lesioni del sistema vestibolare e l‘atassia cerebellare da lesioni del cervelletto.
5. I nuclei vestibolari sono collegati con il vestibolocervelletto o vestibolocerebello. Il vestibolocervelletto è la parte mediana del cervelletto e comprende la corteccia del verme, i nuclei del fastigio e il flocculo. Questa parte del cervelletto corrisponde al lobo flocculo-nodulare ed è quindi completamente sovrapponibile all’archicerebellum, cioè alla parte filogeneticamente più antica del cervelletto.
È connesso ai nuclei vestibolari da cui riceve afferenze dirette provenienti dalle creste ampollari e dalle macule. Queste afferenze raggiungono la corteccia cerebellare sotto forma di fibre muscoidi (fibre muscoidi vestibolo-cerebellari primarie). Inoltre è collegato alla formazione reticolare e ai nuclei basilari del ponte. Le principali efferenze sono dirette ai nuclei vestibolari e costituiscono la prima tappa della via cerebello-vestibolo-spinale. Per queste connessioni il vestibolocerebello è coinvolto nel mantenimento e ripristino dell’equilibrio nella stazione e nella locomozione e nella coordinazione del movimento di testa e occhi.
