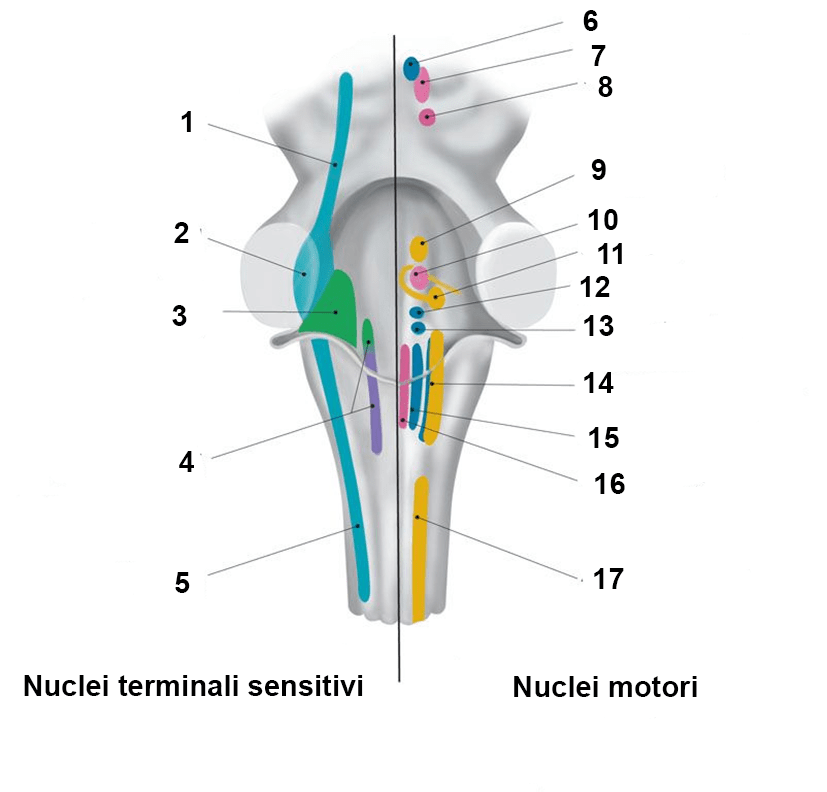
Nuclei terminali sensitivi del n. trigemino (V)
Il n. trigemino è il principale nervo sensitivo della testa. Attraverso le sue tre branche principali (n. olftalmico, n. mascellare e n. mandibolare – che hanno dimensioni pressoché simili, fatto da cui deriva il nome del nervo, cioè costituito da tre parti gemelle) fornisce fibre sensitive somatiche ai tessuti superficiali della testa (cute, sottocute), alle mucose oronasali, ai denti, alla dura madre e ai principali vasi sanguigni intracranici ed è pertanto responsabile della sensibilità generale della testa. Queste fibre derivano dal ganglio sensitivo del V, il più grosso ganglio associato ai nervi cranici. I neuroni sensitivi primari (neuroni di I ordine) che costituiscono il ganglio sono neuroni pseudounipolari (a T) il cui unico prolungamento si divide dopo un breve tratto in una branca periferica, che va a innervare le strutture anatomiche elencate sopra, e una branca centrale che raggiunge il ponte e va a fare sinapsi con i neuroni di II ordine dei nuclei pontino e spinale, due dei tre nuclei terminali sensitivi del nervo. Al nucleo pontino (n. 2 nella figura – più sviluppato e perciò anche detto nucleo principale) arrivano le fibre che conducono stimoli tattili e propriocettivi provenienti dai mm. mimici, mentre al nucleo spinale (n. 3 nella figura) arrivano le fibre che conducono stimoli termici non dolorosi e stimoli dolorifici. Il nucleo spinale, che ha struttura laminare come la sostanza grigia del midollo spinale (di fatto si continua nel corno dorsale del midollo spinale, riceve segnali nocicettivi non solo dall’intero campo sensitivo trigeminale, ma anche dall’orofaringe (n. glossofaringeo), dalla laringofaringe e dalla laringe (n. vago) e dai rami dorsali dei primi nervi cervicali. I nuclei pontino e spinale proiettano, a loro volta, alla formazione reticolare (integrazione sensitivo-motoria) e al talamo controlaterale (sensibilità somatica).
A questi due nuclei se ne deve aggiungere un terzo, il nucleo mesencefalico che, però non è un vero nucleo terminale sensitivo, ma un ganglio che, per motivi tuttora non chiari, è rimasto inglobato all’interno del SNC. A riprova di ciò, i neuroni che lo costituiscono sono anch’essi neuroni pseudounipolari (mentre i neuroni dei nuclei pontino e spinale sono multipolari) di primo ordine. I prolungamenti periferici di questi neuroni decorrono per un certo tratto all’interno del tronco cerebrale e quindi si associano alle altre fibre che costituiscono il V, passano nel n. mascellare e nel n. mandibolare e si distribuiscono ai legamenti periodontali e ai fusi neuro-muscolari dei mm. masticatori (fibre propriocettive). I processi centrali raggiungono il centro della masticazione della formazione reticolare (nucleo sopratrigeminale) che genera gli schemi motori durante la masticazione. Questi schemi consentono la contrazione/decontrazione coordinata dei mm. masticatori: gli assoni dei neuroni del nucleo sopratrigeminale vanno infatti ad innervare i neuroni del nucleo motore del trigemino da cui originano le fibre motrici somatiche (motrici viscerali speciali) per questi muscoli. In questo modo, una volta iniziata la masticazione, pur essendo un atto volontario, prosegue in maniera automatizzata senza che sia necessario un intervento dei circuiti corticali.
Ricorda che abbiamo parlato delle vie sensitive del n. trigemino anche a proposito della via della colonna dorsale-lemnisco mediale, cui si associano fibre provenienti dai neuroni del nucleo pontino che vanno a costituire il lemnisco trigeminale, e della via spino-talamica, cui si associano altre fibre provenienti dai neuroni del nucleo spinale che entrano pure esse nella costituzione del lemnisco trigeminale.

Se vuoi saperne di più e capire meglio scorri la pagina verso il basso
I nervi cranici
Delle dodici paia di nervi cranici, dieci emergono dal tronco cerebrale. Più precisamente due paia dal mesencefalo (III, IV), quattro dal ponte (V-VIII) e quattro dal midollo allungato (IX-XII). Il primo paio (I- n. olfattivo) si stacca dai bulbi olfattivi del telencefalo, mentre il II paio (n. ottico) è associato al diencefalo.
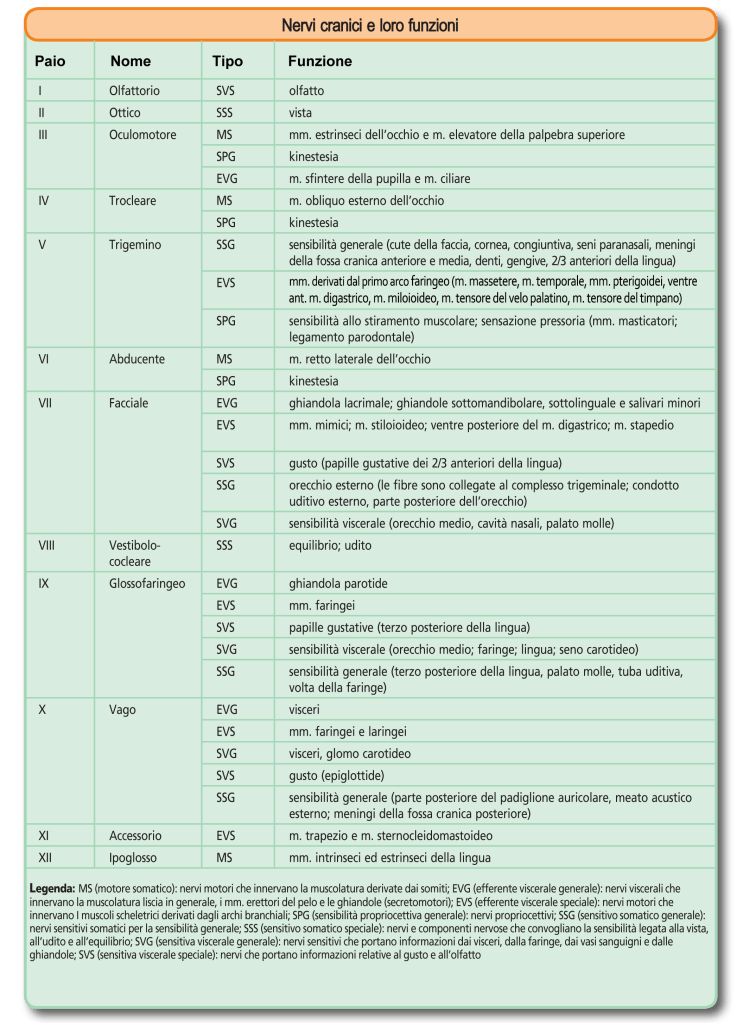
Nella tabella, le fibre che compongono di ciascun nervo sono classificate in tipi diversi secondo le caratteristiche degli organi innervati e/o la loro funzione. Le fibre motrici somatiche (MS) innervano i muscoli striati, volontari, derivati dai somiti. Quelle motrici viscerali speciali (EVS) innervano i muscoli derivati dagli archi branchiali. Occorre fare attenzione alla denominazione di questi diversi tipi di fibre: benché esse innervino muscoli striati e, quindi, volontari, in diversi casi questi muscoli non intervengono, in senso stretto, in quelle che sono le funzioni tipiche dell’apparato locomotore, cioè il mantenimento della stazione quadrupedale e la locomozione. Un’eccezione è in questo senso rappresentata dalle fibre EVS del n. accessorio spinale che sono destinate ad alcuni mm. del collo. A sottolineare le caratteristiche dei muscoli innervati (muscolatura striata volontaria), in diversi testi le fibre EVS sono comunque considerate fibre MS.
Esaminando la tabella è, infatti, possibile vedere che le fibre EVS del n. trigemino si distribuiscono a muscoli quali i mm. masticatori* e il m. miloioideo, che intervengono nella masticazione e quindi, svolgono una funzione che rientra tra quelle dell’apparato digerente; altre fibre EVS che derivano dal V innervano il m. tensore del velo palatino, che, in caso di lesione, può provocare alcune forme di sordità, e il m. tensore del timpano, che riduce i suoni associati alla masticazione. In modo simile le fibre EVS del n. facciale innervano muscoli che non intervengono funzionalmente nella locomozione, ma sono responsabili della mimica facciale (mm. mimici delle labbra e delle guance), o coadiuvano la masticazione/deglutizione (ventre posteriore del m. digastrico* e m. stiloioideo). Infine, il m. stapedio previene un eccessivo movimento della staffa, coadiuvando la trasduzione delle onde sonore in vibrazioni meccaniche nell’orecchio medio. Da ultimo le fibre EVS del n. glossofaringeo e del n. vago si distribuiscono ai mm. faringei (IX e X) e laringei (X) che, rispettivamente, intervengono nella deglutizione e nella fonazione.
*in alcuni testi il m. digastrico è considerato a se stante e non facente parte dei mm. masticatori. Si tenga anche presente che il muscolo è effettivamente costituito da due ventri muscolari nel gatto (come nell’uomo), mentre nel cane esiste un solo ventre muscolare.
Una terza categoria di fibre motrici riscontrabili in alcuni nervi cranici è costituita da fibre efferenti viscerali generali (EVG). Esse sono fibre pregangliari parasimpatiche che derivano dai nuclei motori parasimpatici di quattro diversi nervi cranici: il n. oculomotore, il n. intermedio (una delle due componenti del n. intermedio-facciale o, semplicemente, n. facciale), il n. glossofaringeo e il n. vago. Questi nuclei si chiamano anche nucleo di Edinger-Westphal (III), nucleo salivatorio craniale (VII), nucleo salivatorio caudale (IX) e nucleo motore dorsale del vago (X).
Le fibre sensitive sono classificabili in cinque (o quattro – vedi sotto) tipi diversi. Le fibre sensitive somatiche generali (SSG) raccolgono informazioni riferibili alla sensibilità somatica generale (tattile, termica non dolorifica e dolorifica) degli organi dell’apparato tegumentario e dell’apparato locomotore. Le fibre sensitive somatiche propriocettive trasmettono informazioni relative alla sensibilità propriocettiva generale (SPG), sono quindi responsabili della propriocezione, cioè della raccolta di informazioni sullo stato e la velocità di contrazione dei muscoli striati, sulle forze di trazione esercitate sui tendini e sulle variazioni di ampiezza degli angoli articolari. Si tenga presente che, spesso, il termine kinestesia (cinestesia) è utilizzato come sinonimo di propriocezione e che è possibile fare rientrare anche le fibre propriocettive fra le SSG. Si osservi come tutti i nervi cranici che regolano la contrazione dei mm. oculari estrinseci (III, IV e VI) contengano anche fibre propriocettive. Spesso queste ultime non sono considerate nelle descrizioni di tali nervi per i quali si considerano, per semplicità le sole fibre motrici somatiche o viscerali. Le fibre sensitive somatiche speciali (SSS) raccolgono informazioni relative alla cosiddetta sensibilità somatica speciale (vista, udito, equilibrio). Anche le fibre sensitive viscerali possono essere di tipo generale (SVG) o speciale (SVS). Nel primo caso ricevono stimoli riguardanti la sensibilità generale dei visceri, nel secondo caso stimoli olfattivi o gustativi.
L’esistenza dei diversi tipi di fibre motrici e sensitive a costituire i singoli nervi cranici fa si che i neuroni di origine delle fibre motrici e quelli che ricevono le informazioni sensitive (neuroni di II ordine dei nuclei terminali sensitivi) possano essere raggruppati in colonne di tipo funzionale denominate in maniera corrispondente a quella vista sopra per le fibre che formano i singoli nervi cranici (es. colonna motrice somatica, colonna sensitiva somatica generale, ecc.). Mentre nel midollo spinale le diverse colonne di neuroni sono continue a formare le corna della sostanza grigia, nel tronco cerebrale queste colonne sono frammentate per la complessità dello sviluppo embrionale di questa parte del SNC. Nella figura a inizio pagina sono indicati con colori diversi i nuclei che costituiscono le diverse colonne neuronali del tronco cerebrale (colonna SSG/SPG: verde chiaro; colonna SSS/SVS: verde scuro; colonna SVG: viola; colonna MS: rosa; colonna EVG: blu; colonna EVS: giallo).
Prova a rispondere a queste domande:
- Cosa si intende per colonna della sostanza grigia?
- Nel midollo spinale esiste una colonna efferente viscerale generale?
- Quali sono i recettori che raccolgono le informazioni propriocettive?
- Perchè è importante il nucleo del tratto solitario?
- Che funzione hanno i nuclei cocleari?
(le risposte sono al fondo della pagina)

Ricordati di consultare un libro o cercare informazioni in rete se non conosci il significato di alcuni termini!
Risposte
1. Una colonna della sostanza grigia è un insieme di neuroni che si estende in direzione rostro-caudale nel tronco cerebrale o nel midollo spinale. Si tratta di un termine strettamente correlato alle modalità di sviluppo del primitivo tubo neurale (vedi Embriologia) in relazione alle diverse modalità di proliferazione di tre diverse zone (dall’interno all’esterno: ependimale, mantellare e marginale) riconoscibili in sezioni trasversali del tubo neurale.
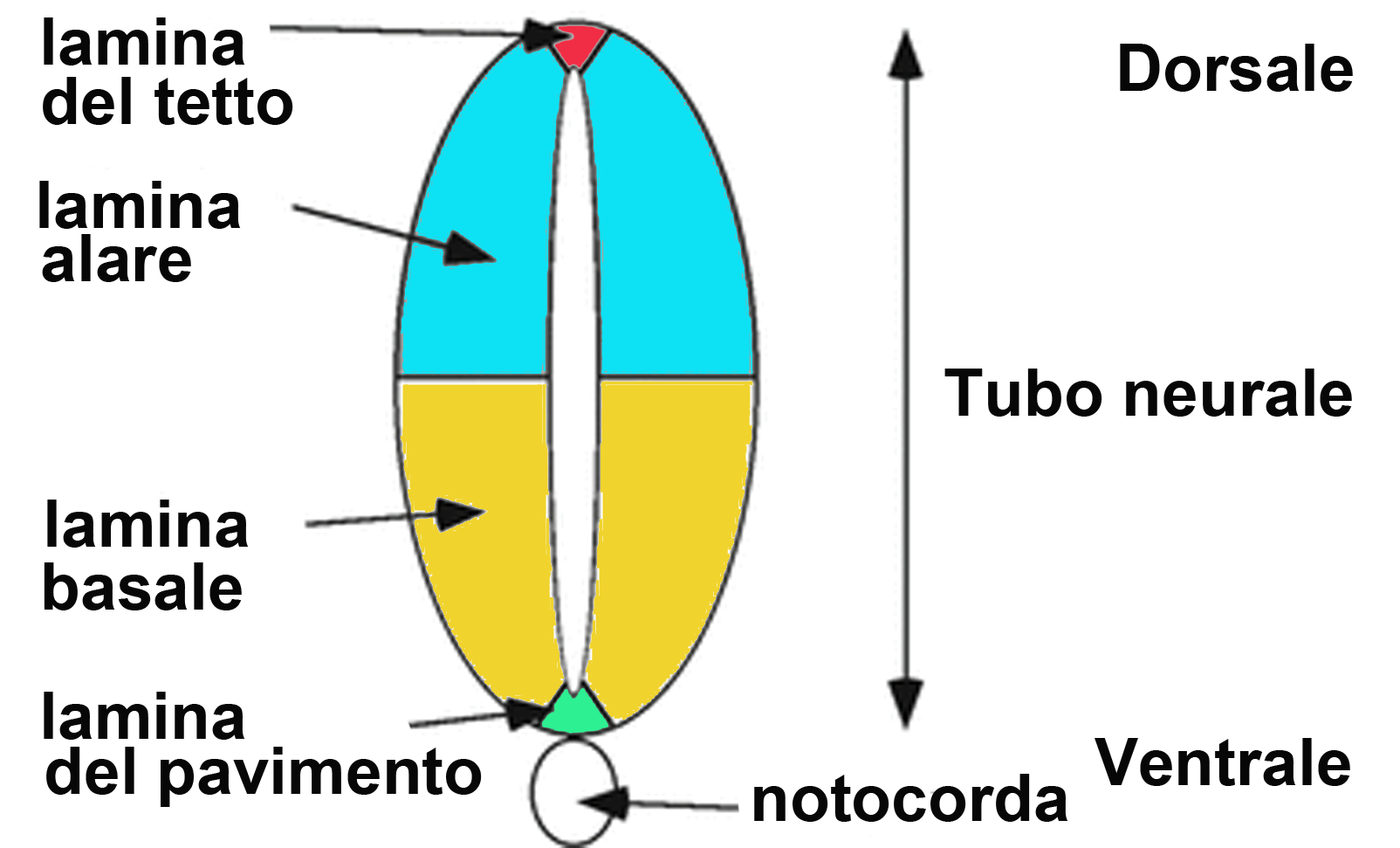
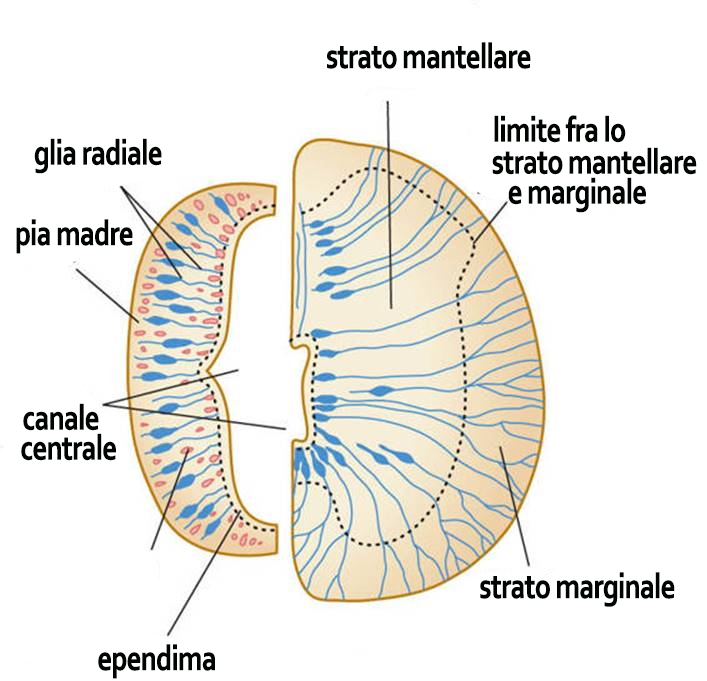
Suddivisione del tubo neurale in lamine (sin) e strati del tubo neurale (dx)
Nel midollo spinale, a sviluppo ultimato, le colonne costituiscono le corna della sostanza grigia, mentre nel tronco cerebrale si frammentano per il passaggio delle fibre nervose che lo raggiungono, lo attraversano o lo abbandonano. In questo modo si formano nuclei di neuroni tra loro separati.
2. Esiste una colonna efferente viscerale generale (EVG) che si presenta con aspetti diversi secondo le diverse parti del midollo spinale. La colonna corrisponde alla sostanza grigia intermediolaterale che unisce tra loro le basi delle corna dorsali e ventrali. A livello del midollo spinale toraco-lombare la colonna EVG è ben evidente e forma un vero e proprio corno laterale della sostanza grigia. In esso sono localizzati i corpi cellulari dei neuroni pregangliari del sistema simpatico. A livello del midollo spinale sacrale il corno laterale non è molto evidente e i neuroni pregangliari del sistema parasimpatico sono raggruppati nella sostanza grigia intermediolaterale.
3. I recettori che raccolgono informazioni propriocettive sono i fusi neuro-muscolari, gli organi tendinei di Golgi e i meccanocettori delle articolazioni. I fusi neuro-muscolari raccolgono informazioni sull’intensità e velocità di contrazione delle fibre muscolari extrafusali. Gli organi tendinei di Golgi registrano la trazione esercitata sui tendini. I meccanocettori articolari sono meccanocettori a bassa soglia (corpuscoli di Pacini e di Ruffini) che monitorano lo stato di apertura/chiusura deli angoli articolari. I corpi cellulari di origine delle terminazioni che costituiscono questi recettori si trovano nei gangli spinali (tutti i tipi recettoriali), nel ganglio (meccanocettori articolari) o nel nucleo mesencefalico del trigemino (fusi neuromuscolari e organi tendinei di Golgi).
4. Il nucleo del tratto solitario (NST) è uno dei principali nuclei sensitivi del midollo allungato dorsale, che riceve informazioni cardiovascolari, viscerali, respiratorie, gustative e oro tattili. Si estende da un livello corrispondente alla decussazione delle piramidi, ai limiti con il midollo spinale cervicale, rostralmente fino al terzo caudale del nucleo cocleare dorsale. Caudalmente, l’NST attraversa la linea mediana formando i subnuclei commissurali. Rostralmente all’area postrema, l’NST si divide in due metà che circondano la linea mediana e la sommità ventro-laterale del IV ventricolo. Sull’NTS convergono praticamente tutte le informazioni sensitive viscerali che sono veicolate al SNC attraverso i nervi cranici. Ricorda che l’NTS può essere considerato parte della formazione reticolare. Infatti in esso si trovano il centro cardiovascolare, l’area chemorecettrice (che si estende anche ai subnuclei commessurali), i centri vasomotori e parte del centro della respirazione della formazione reticolare che controllano, rispettivamente, l’attività cardiaca, il pH del sangue e del liquor, la pressione sanguigna e l’attività respiratoria.
5. I nuclei cocleari sono i nuclei sensitivi terminali del n. cocleare (VIIIc) che ricevono informazioni dalle fibre sensitive somatiche speciali che originano dai neuroni del ganglio cocleare. I nuclei cocleare sono la prima stazione delle vie acustiche centrali che hanno il compito di trasferire le informazioni sonore alla corteccia uditiva del lobo temporale dell’emisfero cerebrale dove diventeranno coscienti.
Ricorda che il suono inteso come onde di rarefazione e compressione del mezzo è stato trasformato in segnale elettrico dalla coclea, dove avviene una trasduzione meccano-elettrica, cioè la trasformazione del suono in un segnale elettrico. Pertanto quando si parla di vie acustiche centrali non dobbiamo più pensare agli stimoli sonori come onde sonore ma solo come un segnali elettrici. Le fibre nervose provenienti da un orecchio si portano ad entrambi i nuclei cocleari dei due lati del ponte: in questo modo avviene un fenomeno importante detto fusione binaurale nel quale interviene un insieme di nuclei di neuroni che formano il corpo trapezoide. Gli animali ascoltano i suoni con entrambe le orecchie e le percezioni sonore avvengono separatamente, però al cervello giunge un solo suono, proprio grazie a questo meccanismo di fusione. Il nuovo segnale è, per così dire, una creazione del cervello che non esiste nella realtà. Questo è un esempio tipico di quelli che vengono definiti processi di integrazione degli stimoli a livello centrale.
I nuclei cocleari sono due, il nucleo cocleare dorsale e il nucleo cocleare ventrale.
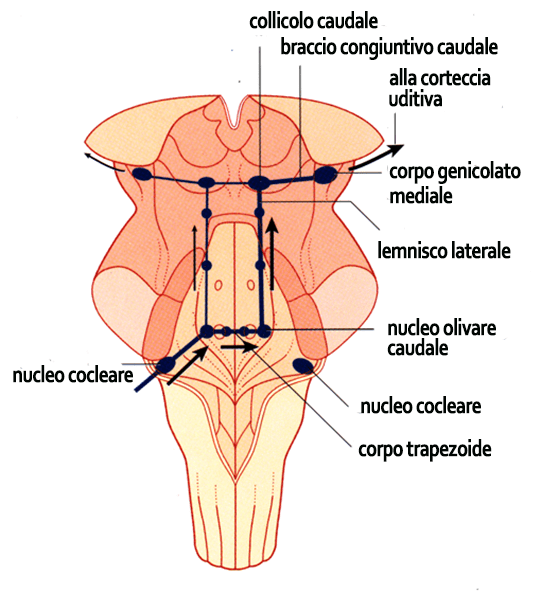
Sempre nel ponte si trova il nucleo olivare craniale. I nuclei olivari craniali si dividono in due parti su ciascun lato: laterale e mediale. Interessante è l’azione del parte laterale del nucleo olivare che è in grado di percepire i diversi caratteri del suono, soprattutto la differenza d’intensità. Analogo interesse riveste la parte mediale che è in grado di analizzare le differenze temporali con cui il suono ha raggiunto le due orecchie. Grazie a queste due azioni il cervello degli animali è in grado di localizzare la provenienza di un suono.
Ipotizziamo che un suono provenga da destra, questo raggiunge prima l’orecchio destro e poi il sinistro (analisi temporale svolta dal nucleo olivare craniale mediale). Inoltre le onde sonore a destra saranno un poco più intense rispetto a quelle che raggiungono l’orecchio sinistro, questo perché l’orecchio sinistro risente dell’azione mascherante della testa. Questa differenza d’intensità viene analizzata dal nucleo olivare craniale laterale. La capacità di localizzare la provenienza di un suono è una delle funzioni principali dell’ascolto binaurale.
A questo punto il segnale elettrico sonoro percorre la via del lemnisco laterale. I lemnischi laterali (che non sono altro che piccoli fasci di fibre nervose) sono ben distinti a destra e sinistra e non ci sono fibre di interconnessione. Essi arrivano al collicolo caudale. I due collicoli caudali (destro e sinistro) hanno la funzione di integrare le informazioni provenienti dal nucleo cocleare dorsale e dal complesso olivare.
Dal collicolo caudale alcune fibre che trasportano le informazioni sonore arrivano al talamo, da dove partono fibre per attivare la corteccia cerebrale, integrando i segnali sonori con quelli di altra natura. Quindi il talamo svolge una funzione associativa tra diverse aree corticali. Altre fibre raggiungono il corpo genicolato mediale. Quest’ultimo è un nucleo che si trova nella parte posteriore del talamo. Dai corpi genicolati mediali partono le fibre nervose che raggiungono la corteccia uditiva. Viene chiamata corteccia uditiva primaria la zona dove arrivano direttamente i segnali elettrici provenienti dalle vie acustiche.
