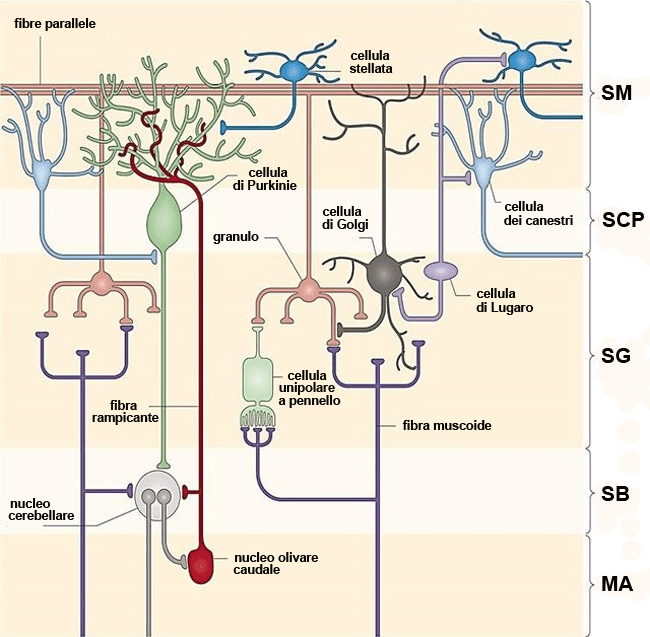
Fibre parallele
Le fibre parallele dello strato molecolare sono gli assoni dei granuli cerebellari. Ciascuno di essi corre verso la superficie della corteccia del cervelletto e si divide a “T” lungo l’asse delle singole lamine cerebellari, incrociando le arborizzazioni dendritiche di molte cellule di Purkinje, con cui forma sinapsi eccitatorie “en-passant”. In ciascuna lamina cerebellare si trovano milioni di fibre parallele. Diverse centinaia di migliaia di fibre parallele fanno sinapsi con (ed eccitano) ciascuna cellula di Purkinje. Gli assoni delle fibre parallele sono anche il principale input eccitatorio per le cellule di Golgi, stellate e dei canestri. Ogni fibra parallela fornisce un piccolo input sinaptico a ciascuna cellula di Purkinje, ma i segnali in ingresso di molte fibre parallele possono sommarsi fra loro per attivare le cellule di Purkinje.

Se vuoi saperne di più e capire meglio scorri la pagina verso il basso
I circuiti cerebellari
I circuiti della corteccia cerebellare sono abbastanza ben caratterizzati (vedi la figura a inizio pagina). Esistono solo due tipi di assoni che entrano nel cervelletto, le fibre muscoidi e le fibre rampicanti. Entrambi questi tipi di fibre rilasciano neurotrasmettitori eccitatori, glutamato nel caso delle fibre muscoidi e aspartato nel caso delle fibre rampicanti. Quando entrano nel cervelletto, sia le fibre muscoidi che quelle rampicanti hanno rami collaterali che fanno sinapsi (ed eccitano) i neuroni dei nuclei cerebellari profondi. Tuttavia, questi due tipi di fibre hanno terminazioni ed effetti abbastanza diversi nella corteccia cerebellare.
Le fibre muscoidi si ramificano ampiamente quando entrano nella corteccia. Ogni loro terminale fa sinapsi su un gruppo di dendriti delle cellule dei granuli, eccitandole. Gli assoni dei granuli formano le fibre parallele che entrano in contatto sinaptico con le cellule di Purkinje e gli interneuroni inibitori dello strato molecolare (vedi sopra). Gli assoni delle cellule di Purkinje sono le uniche fibre nervose a lasciare effettivamente la corteccia cerebellare e a fare sinapsi sui neuroni dei nuclei cerebellari e su alcuni neuroni vestibolari, inibendoli potentemente. Pertanto, l’input eccitatorio che entra nel cervelletto con la fibra muscoide è prima responsabile dell’eccitazione dei neuroni dei nuclei profondi e quindi dell’eccitazione dei granuli che, a loro volta stimolano le cellule di Purkinje, le quali “spengono” il neuroni dei nuclei profondi. Si viene quindi a creare quello che è un “circuito di base” che regola i movimenti acquisiti:
Fibre muscoidi→Granuli→Neuroni di Purkinje→Neuroni nuclei centrali
È fondamentale che una quantità corretta di inibizione arrivi ai nuclei cerebellari profondi al fine di produrre un output appropriato, inibendo l’attività indesiderata nei neuroni di questi nuclei. Questo è un processo simile alla scultura, in cui le parti indesiderate della pietra vengono rimosse per creare un motivo.
Le fibre rampicanti cerebellari fanno invece sinapsi direttamente sulle cellule di Purkinje e le eccitano con forza. C’è solo una fibra rampicante per cellula di Purkinje e ogni fibra rampicante contatta solo da una a tre cellule di Purkinje. Pertanto, esiste un’intima connessione tra i neuroni del nucleo olivare caudale e le cellule di Purkinje che è possibile schematizzare nel circuito che segue:
Fibre rampicanti→Neuroni di Purkinje→Neuroni nuclei centrali
Il nucleo olivare caudale è un “rilevatore di errori” che si attiva durante l’apprendimento motorio, quando l’animale compie degli errori mentre impara un nuovo movimento o affronta una situazione che comporta la necessità di modificare i parametri di un movimento noto. Quando una particolare azione va fuori bersaglio, i neuroni del nucleo olivare caudale vengono attivati. Ciò si traduce in una potente attivazione delle cellule di Purkinje attraverso le fibre rampicanti. Questa potente attivazione delle cellule di Purkinje inibisce i neuroni dei nuclei cerebellare profondi, sperando di porre fine alla componente indesiderata dell’azione.
Se è così che funziona il cervelletto, in che modo il cervelletto contribuisce all’apprendimento motorio, in modo che la prossima volta che viene eseguito il movimento, venga eseguito in modo più accurato dall’inizio? Questo sembra essere il risultato della plasticità della sinapsi tra la fibra parallela e la cellula di Purkinje. Al momento dell’attivazione della cellula di Purkinje da parte della fibra rampicante, tutte le sinapsi delle fibre parallele recentemente attive subiranno un processo di depressione a lungo termine, cioè vengono inibite per un lasso di tempo relativamente lungo (alcune ore). Ciò produce una depressione a lungo termine delle risposte successive all’input attraverso quella particolare fibra parallela che era attiva immediatamente prima che si attivasse la fibra rampicante, vale a dire che le sinapsi che erano attive nel momento dell’input della fibra rampicante saranno indebolite, in modo che la prossima volta che la fibra parallela specifica sarà attiva, avrà un effetto eccitatorio minore sulla cellula di Purkinje.
Poiché il nostro attuale concetto di funzione delle fibre rampicanti è che esse trasmettano un segnale di errore, le sinapsi delle cellule dei granuli alle cellule di Purkinje che erano attive al momento dell’errore saranno inibite. Ciò è appropriato poiché queste sinapsi erano probabilmente proprio quelle responsabili dell’attività indesiderata. Pertanto, ciascuna sinapsi può essere regolata durante un processo di apprendimento per produrre il corretto output cerebellare. Nel caso di schemi motori, ciò consente l’apprendimento procedurale, dove ogni volta che viene eseguita un’azione, essa diventa un pò più accurata poiché le “sinapsi giuste” contribuiscono alla risposta.
Meno si sa sull’attività degli altri neuroni della corteccia cerebellare. È interessante notare che tutti gli altri tre tipi di cellule (Golgi, stellate e dei canestri) sono inibitori. Ciascuno di questi tipi di cellule è eccitato dalle fibre parallele e ciascuna si collega su una certa parte del circuito (vedi figura a inizio pagina). Le cellule di Golgi hanno un effetto inibitorio retroattivo (feedback) sulle cellule dei granuli. L’attivazione delle cellule stellate da parte delle fibre parallele ha invece un effetto di controllo che porta a inibire le cellule di Purkinje poco dopo che esse stesse sono eccitate dalle fibre parallele. Questa inibizione può “spegnere” l’attività delle cellule di Purkinje subito dopo che sono state eccitate, consentendo alla cellula di Purkinje di rispondere solo all’attività immediata nei granuli e nelle fibre parallele. Le cellule dei canestri sono un caso speciale, poiché i loro assoni di solito raggiungono le cellule di Purkinje circostanti, non quelle a cui sono adiacenti. Ciò può “disattivare” l’attività nelle cellule di Purkinje a una certa distanza attorno al centro dell’attività. Sebbene esistano diverse idee sul funzionamento di questi neuroni inibitori della corteccia cerebellare, è doveroso dire che tutti i dettagli non sono compresi e, nonostante la sua architettura piuttosto semplice, non abbiamo modelli computerizzati funzionanti dei circuiti che possano simulare le attività complesse che il cervelletto regola così efficacemente. È chiaro, tuttavia, che i danni ai nuclei cerebellari o alle cellule di Purkinje hanno effetti gravi e potenzialmente devastanti da un punto di vista clinico.
Prova a rispondere a queste domande:
- Sai citare alcuni movimenti regolati da schemi motori?
- Quale neurotrasmettitore utilizzano le cellule di Purkinje?
- Il cervelletto è collegato direttamente con la sostanza reticolare?
- Il cervelletto è collegato direttamente con il talamo?
- Il cervelletto è collegato direttamente con la corteccia cerebrale?
(le risposte sono al fondo della pagina)

Ricordati di consultare un libro o cercare informazioni in rete se non conosci il significato di alcuni termini!
Risposte
1. La masticazione, la deglutizione, la locomozione sono movimenti regolati da schemi motori. Questi movimenti sono acquisiti per apprendimento procedurale durante le prime fasi della vita postnatale e poi diventano automatici di modo che l’animale può eseguirli senza che sia necessario un controllo diretto da parte della corteccia cerebrale.
2. Le cellule di Purkinje sono neuroni inibitori e, quindi, utilizzano il GABA come neurotrasmettitore.
3. Si, attraverso il fascio cerebello-reticolare i cui assoni originano dal nucleo interposito dello spinocerebello. Il fascio cerebello-reticolare regola l’attività dei fasci reticolo-spinali che intervengono nel controllo della postura e della locomozione.
4. Si attraverso il fascio dento-talamico che origina dai neuroni del nucleo dentato e raggiunge il talamo e, di qui, la corteccia motrice. Si tratta di un collegamento che interviene nella programmazione del movimento.
5. No.
