Lobo temporale dell’emisfero cerebrale
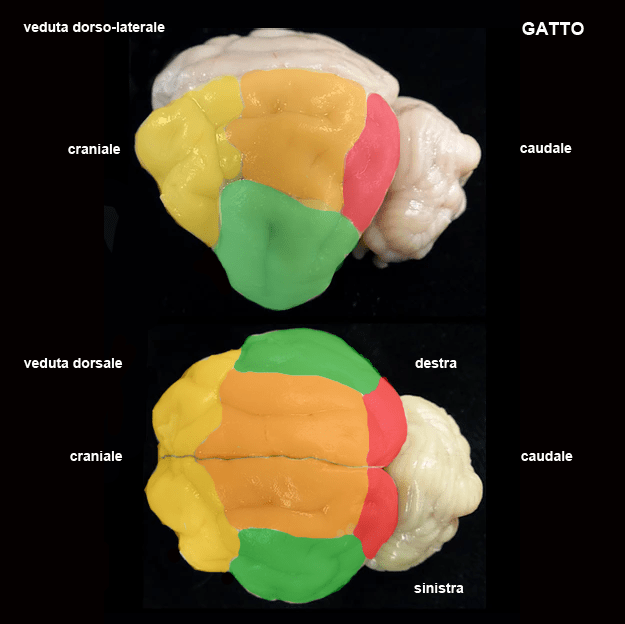
Le immagini mostrano la suddivisione in lobi degli emisferi cerebrali. Il lobo temporale è colorato in verde, il lobo frontale in giallo, il lobo parietale in arancione e il lobo occipitale in rosso. Nell’immagine non sono visibili il lobo olfattivo e il lobo limbico. Pertanto negli animali di primario interesse per la Medicina Veterinaria in ciascun emisfero cerebrale sono riconoscibili sei lobi, a differenza dell’uomo dove il lobo olfattivo è pressoché inesistente. Nelle specie in cui le funzioni olfattive sono molto sviluppate è visibile sulla faccia ventrale di ciascun emisfero una zona della corteccia cerebrale che si presenta a superficie pressoché liscia. Essa è chiamata in alcuni testi lobo piriforme, ma dovrebbe essere più propriamente detta corteccia piriforme.

Se vuoi saperne di più e capire meglio scorri la pagina verso il basso
Emisferi cerebrali
Gli emisferi cerebrali costituiscono la parte più sviluppata del telencefalo. Sono rivestiti dalla corteccia cerebrale (pallio), uno strato di sostanza grigia che si localizza alla superficie degli emisferi dopo una serie di movimenti migratori nel corso dello sviluppo e della neurogenesi. Si tratta di una serie di morfogenetici molto complessa che porta anche alla formazione dell’ippocampo, nella parte profonda di ciascun emisfero. Al di sotto della corteccia cerebrale, ciascun emisfero cerebrale è costituito da una voluminosa massa di sostanza bianca nella quale sono inglobati una serie di nuclei di sostanza grigia (nuclei della base) che, nell’insieme, formano il corpo striato. Gli emisferi cerebrali formano la parte più sviluppata del cervello, che comprende anche il diencefalo e le formazioni commessurali.
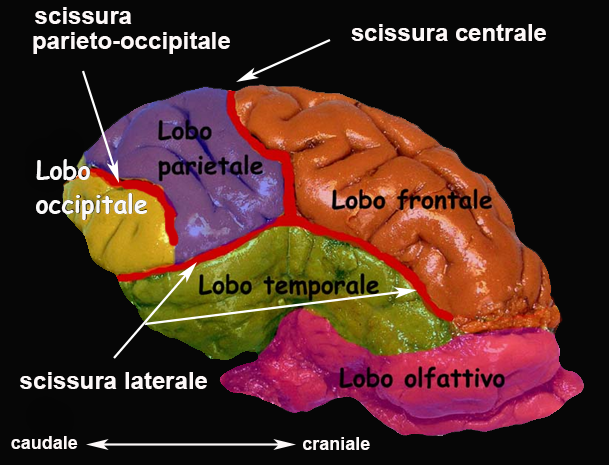
Le scissure e i solchi presenti sulla superficie corticale consentono di suddividere ciascun emisfero in lobi: frontale e parietale (rispettivamente al davanti e al di dietro della scissura centrale o del solco crociato in alcune specie di animali domestici), temporale (al di sotto della scissura laterale), occipitale (situato posteriormente alla scissura parieto-occipitale). Profondamente alla scissura laterale si trova l’insula, mentre a circondare il corpo calloso sulla faccia mediale dell’emisfero è presente la corteccia cingolata che costituisce il lobo limbico dell’emisfero cerebrale.

Corteccia cerebrale
La corteccia cerebrale costituisce la porzione gerarchicamente più importante del telencefalo. Alla posizione corrisponde un’analoga attitudine funzionale: si ritiene infatti che la corteccia cerebrale sia responsabile, nell’uomo, delle cosiddette funzioni cerebrali superiori (linguaggio, pensiero astratto, comportamento sociale, apprendimento, ecc.). La sua importanza funzionale è testimoniata anche dall’aumento relativo della sua superficie che si realizza nel corso della filogenesi, fino ad assumere, nei Primati non umani e poi in particolare nell’uomo, il massimo sviluppo. Per accomodarne l’estensione all’interno del cranio, il mantello corticale è fortemente ripiegato a formare scissure e solchi che si approfondano nella sottostante sostanza bianca. Tra i solchi e le scissure, dove sono decorrono i vasi encefalici superficiali, sono comprese le circonvoluzioni (o giri), visibili sulla superficie esterna dell’encefalo. In base alla presenza o meno di scissure e solchi superficiali e, conseguentemente, di circonvoluzioni cerebrali, gli animali possono essere suddivisi in girencefali e lissencefali. Gran parte degli animali che interessano la Medicina Veterinaria clinica appartengono alla prima categoria, con l’eccezione del coniglio che è un lissencefalo, come gran parte dei Roditori da laboratorio.
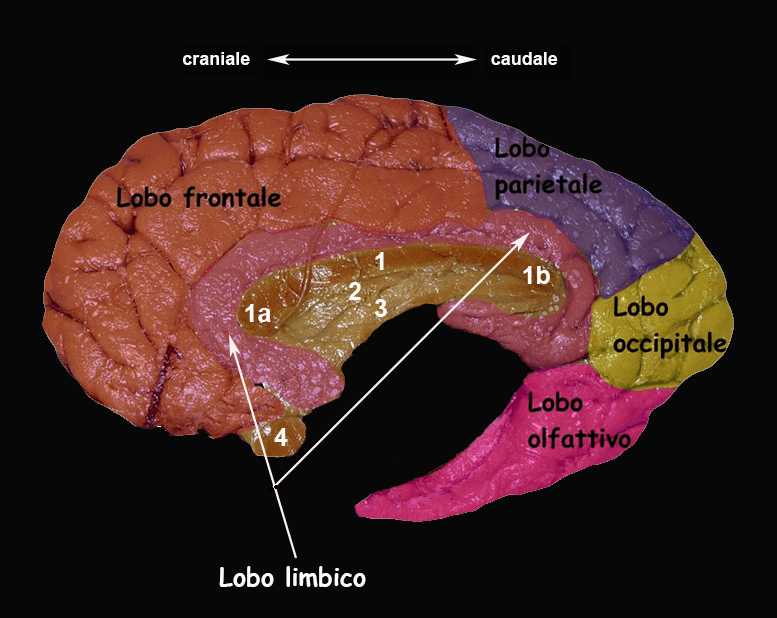
Su base filogenetica è possibile suddividere la corteccia cerebrale in tre porzioni: la paleocortex (o paleopallio), rappresentata dalla corteccia olfattiva; l’archicortex (o archipallio), che comprende l’ippocampo; la neocortex (o neopallio), che costituisce la maggior estensione della corteccia. Archicortex e paleocortex, costituiscono i sistemi (lobi) limbico e olfattivo. Quest’ultimo è molto sviluppato in diverse specie animali.
La corteccia dei diversi lobi degli emisferi cerebrali presenta caratteristiche diverse, nonché diverse attitudini funzionali. Tuttavia, la distinzione in base a criteri topografici non risulta soddisfacente e si preferisce basarsi su criteri citoarchitettonici, che prendono come riferimento parametri anatomo-microscopici quali ad esempio lo spessore relativo dei diversi strati. Basandosi su questo criterio, è possibile individuare fondamentalmente tre tipi di corteccia cerebrale:
la corteccia agranulare presenta il quarto strato (talamo-recipiente) poco sviluppato e la prevalenza del quinto strato. Si tratta di un tipo di corteccia adatto a mediare le efferenze; ne è un tipico esempio l’area motoria primaria;
la corteccia granulare (o koniocortex) è caratterizzata invece da un notevole sviluppo dello strato talamo-recipiente. E’ quindi è un tipo di corteccia adatto a ricevere le afferenze; ne sono un esempio le aree somatosensitiva, uditiva e visiva primaria;
la corteccia associativa è caratterizzata da un sostanziale equilibrio tra gli strati e i tipi cellulari. Invece che nella ricezione di informazioni sensoriali dal talamo o nell’invio di informazioni ai centri motori, è tipicamente coinvolta in funzioni di integrazione (funzioni superiori). Le connessioni di questo tipo di corteccia sono prevalentemente connessioni cortico-corticali, anche se non mancano afferenze talamiche ed efferenze verso formazioni sottocorticali.
Una suddivisione più precisa della corteccia in senso tangenziale è quella individuata da Brodmann all’inizio del secolo scorso. Questo studioso ha suddiviso il mantello corticale in circa 50 aree numerate con numeri arabi. Questa classificazione, basata su criteri rigorosamente citoarchitettonici, ha un importante corrispettivo funzionale ed è pertanto di notevole utilità, tanto che ad essa ci si riferisce ancora oggi nella maggior parte delle trattazioni sulla corteccia cerebrale. Nel corso degli anni la suddivisione corticale in aree è stata rivista e aggiornata alla luce dei dati sperimentali che si andavano accumulando.
Nonostante struttura e funzione della corteccia cerebrale siano molto complesse e ancora non completamente note, essa è costituita molto semplicemente da due soli tipi di neuroni: le cellule piramidali e le cellule stellate (o dei granuli).
Le cellule piramidali (neuroni piramidali) sono neuroni di proiezione, con un lungo assone, che stabiliscono contatti a distanza con neuroni situati in altre zone corticali o in strutture sottocorticali. I neuroni piramidali presentano una forma assai caratteristica, con un corpo cellulare triangolare, ad apice rivolto verso l’alto. Dalla base del corpo emergono 4-6 dendriti basali, mentre un singolo dendrite apicale di grosso diametro si dirige verso l’alto (cioè verso la superficie piale).
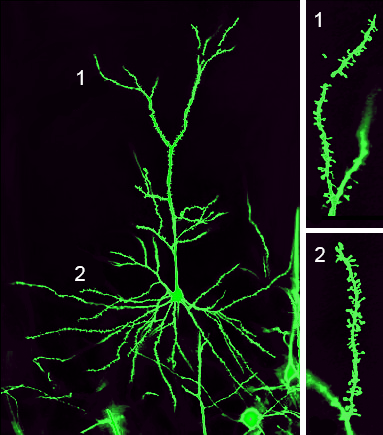
L’assone invece è diretto verso il basso, per raggiungere la sostanza bianca situata nella parte profonda dell’emisfero. Le diramazioni dei dendriti basali e del dendrite apicale sono fittamente ricoperte di spine, sulle quali si stabiliscono numerosissimi contatti sinaptici. Data l’estensione dell’albero dendritico e la lunghezza dell’assone, i neuroni piramidali hanno il compito di integrare un notevole numero di informazioni e trasmetterle a distanza. Rappresentano quindi le cellule di output della corteccia cerebrale. Il loro neurotrasmettitore è il glutamato.
Le cellule stellate hanno come caratteristica comune un’evidente ramificazione dell’assone, che tuttavia è breve e dà vita a terminali i quali rimangono all’interno della corteccia cerebrale stessa, nella maggior parte dei casi addirittura nei dintorni del corpo cellulare di origine. Queste cellule sono pertanto interneuroni o neuroni a circuito locale. Alcuni interneuroni corticali utilizzano il neurotrasmettitore eccitatorio glutamato, mentre la categoria di interneuroni più studiata è costituita da cellule inibitorie, che usano il GABA come neurotrasmettitore.
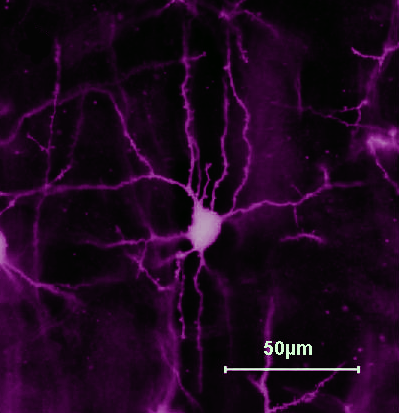
In realtà la semplicità della composizione citologica della corteccia cerebrale non deve ingannare: anche il tipo cellulare più stereotipato (i neuroni piramidali) presenta una notevole disomogeneità nella fine morfologia dell’albero dendritico, così come nelle proprietà fisiologiche, a seconda delle aree corticali considerate. Allo stesso modo, la diversità degli interneuroni GABAergici, nota da tempo, rappresenta probabilmente uno degli aspetti funzionalmente più importanti nella corteccia. Molti degli interneuroni inibitori prendono contatto sinaptico con il soma e la porzione iniziale dell’assone dei neuroni piramidali, influenzandone quindi i segnali in uscita (output); altri interneuroni formano sinapsi sui dendriti dei neuroni piramidali, controllando così, spesso in modo settoriale, i segnali in ingresso verso queste cellule (input). Inoltre, gli interneuroni possono essere ulteriormente suddivisi in base a caratteristiche neurochimiche , al tipo di canali ionici presenti sulla membrana e, di conseguenza, alle loro proprietà elettrofisiologiche. Ne emerge un quadro di grande ricchezza e varietà che, seppure non ancora noto nei dettagli, rappresenta sicuramente la base strutturale della grande complessità fisiologica della corteccia cerebrale.
La corteccia cerebrale è organizzata in strati (vedi Istologia e/o Anatomia microscopica): nel paleopallio e nell’archipallio gli strati sono, rispettivamente, tre o quattro, mentre nel neopallio sono sei. Sebbene la struttura laminare (a strati) della corteccia cerebrale sia nota da molto tempo, il preciso ruolo funzionale che le deve essere attribuito non è ancora definitivamente chiaro. Molte evidenze suggeriscono che la struttura a strati offra un vantaggio computazionale nello svolgimento delle funzioni della corteccia, costituendo un ottimo compromesso tra funzioni analitiche (ad esempio di localizzazione spaziale dello stimolo) e funzioni di sintesi cognitiva (analisi del che cosa lo stimolo rappresenta nel suo insieme).
Per semplicità, da un punto di vista anatomo-funzionale, i sei strati della neocorteccia possono essere ridotti a tre, assumendo che lo strato centrale della circuiteria corticale sia quello talamo-recipiente (il quarto strato, granulare). Gli strati al di sopra del quarto possono essere definiti sopragranulari e, analogamente, possono essere definiti sottogranulari il quinto e sesto strato. Le informazioni vengono trasmesse dalle afferenze talamiche agli interneuroni eccitatori dello strato 4. Da qui, il flusso informativo viene convogliato ai neuroni piramidali degli strati sopragranulari, situati nella stessa colonna corticale. I neuroni degli strati sopragranulari collegano fra loro colonne distanti, nonché, tramite collaterali assonali, trasferiscono informazioni ai neuroni piramidali degli strati infragranulari, che assicurano i collegamenti di ritorno al talamo e alle altre formazioni sottocorticali. Questa descrizione dei microcircuiti della corteccia cerebrale è volutamente semplificata. Si dovrà tenere conto, infatti, del ruolo svolto dagli interneuroni inibitori e della presenza di estese comunicazioni tra colonne vicine.
Prova a rispondere a queste domande:
- Che cos’è un lobo dell’emisfero cerebrale?
- Che cos’è una circonvoluzione cerebrale?
- Che cosa sono la corteccia omotipica e eterotipica?
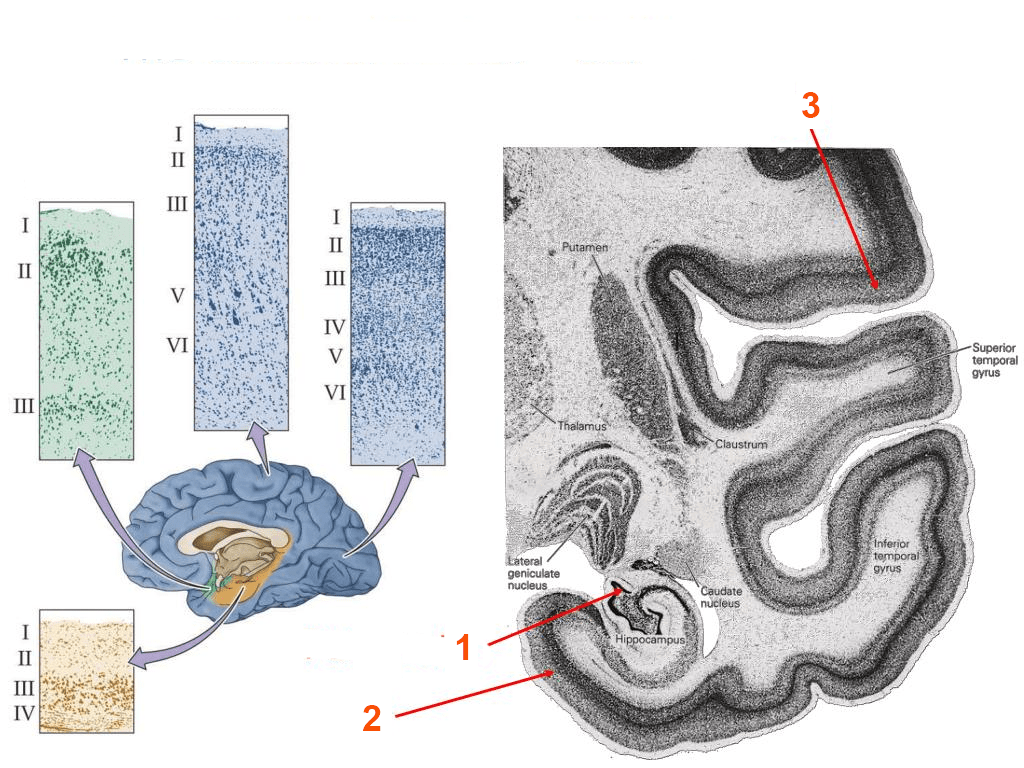
- Nella figura quale parte della corteccia cerebrale indica il numero 2? A quale rappresentazione grafica corrisponde nella parte sinistra dell’immagine?
(le risposte sono al fondo della pagina)
Risposte
1. Un lobo di un emisfero cerebrale è una grossa suddivisione dell’emisfero delimitata da alcuni solchi o scissure. Ciascun lobo comprende diverse circonvoluzioni cerebrali e/o aree corticali. La maggior parte dei lobi degli emisferi cerebrali prende il proprio nome dalle ossa craniche con cui è a contatto indiretto per l’interposizione delle meningi.
2. Una circonvoluzione cerebrale è una parte della corteccia cerebrale delimitata da solchi. Le circonvoluzioni si indicano con il nome del lobo o della regione cui appartengono e con un numero (es. prima circonvoluzione frontale frontale) o precisandone l’orientamento (es. circonvoluzione frontale ascendente); nel caso che limitino una scissura, precisando la loro posizione rispetto alla scissura stessa (es. circonvoluzione ectosilviana); a volte sono indicate con il nome di chi ne ha illustrato il significato funzionale.
3. La corteccia cerebrale omotipica presenta tutti e sei i tipici strati corticali ben sviluppati, mentre la corteccia cerebrale eterotipica non presenta tutti e sei gli strati. Esistono infatti differenze nella struttura della corteccia: regioni diverse hanno spessore ed estensione degli strati diversi e variano per densità e tipi di cellule. Sulla base di queste differenze, si distinguono cinque tipi di corteccia su base citoarchitettonica: agranulare, di tipo frontale, di tipo parietale, di tipo polare e granulare (o koniocorteccia). I tipi corticali frontale, parietale e polare sono costituiti da corteccia omotipica, i tipi agranulare e granulare da corteccia eterotipica. La corteccia cerebrale agranulare è caratterizzata dall’assenza (o dalla scarsissima rilevanza) degli strati 2 e 4 e da evidenti strati 3 e 5 ed è tipica delle aree motorie. La corteccia cerebrale granulare è caratterizzata dall’abbondanza di cellule rotondeggianti, dal notevole sviluppo degli strati 1 e 4 e dallo scarso sviluppo degli strati 3 e 5, caratteristiche queste delle aree sensitive.
4. Nella figura i numeri rappresentano: 1. archipallio la cui struttura tristratificata è schematizzata in verde nel disegno a sinistra; 2. paleopallio, con struttura quadristratificata, in giallo a sinistra; 3. neopallio, a sei strati, in azzurro a sinistra.

Ricordati di consultare un libro o cercare informazioni in rete se non conosci il significato di alcuni termini!
